- 移动端


上海一研生物科技有限公司
10 年
手机商铺
商家活跃:
产品热度:
- NaN
- 0
- 0
- 2
- 2
全部产品
- 查看全部分类
- 实验抗体
- 酶联免疫elisa分析试剂盒
- RNA/DNA提取
- 生化试剂
- 标准品
- ATCC细胞
- 抗体
- 菌种
- elisa检测试剂盒
- 生物抗体
- USP标准品
- 科研抗体
- 原代细胞
- 科研抗体
- 生化试剂类
- PCR检测试剂盒
- 培养基
- 生化检测试剂盒
- 进口品牌
- 代理进口品牌
- 生物碱
- 黄酮
- 查尔酮
- 氧杂蒽酮
- 木脂素
- 香豆素
- 苯丙素
- 其它酚类
- 三萜
- 药物杂质及中间体
- 其它天然产物
- 蒽醌
- 其它醌类
- 甾体
- 二萜
- 其它萜类
- 环烯醚萜
- 倍半萜
- 中药对照品
- 质粒
- 定做种属检测试剂盒/中药材鉴定试剂盒
- 细胞生物学试剂
- DRG试剂盒
- 荧光定量PCR试剂盒
- 对照品
- 菌种库
- 细胞株
- 耗材和仪器
- 分子生物学试剂盒
- 培养基
- ELISA Kit
- ELISA检测试剂盒
- 细胞培养
- 标准溶液
- LAMP试剂盒
- 基因PCR试剂盒
- 鉴定PCR试剂盒
- 进口抗体
- 抑制剂
G蛋白偶联受体66/神经调节肽U受体1抗体
询价
推荐产品
公司新闻/正文
G蛋白偶联受体48抗体验原理
233 人阅读发布时间:2022-03-16 14:22
G蛋白偶联受体48抗体验原理 :
(1)特异性结合抗原:抗体本身不能直接溶解或杀伤带有特异抗原的靶细胞,通常需要补体或吞噬细胞等共同发挥效应以清除病原微生物或导致。然而,抗体可通过与病毒或毒素的特异性结合,直接发挥中和病毒的作用。
(2)活补体:IgM、IgG1、IgG2和IgG3可通过经典途径激活补体,凝聚的IgA、IgG4和IgE可通过替代途径激活补体。
(3)结合细胞:不同类别的免疫球蛋白,可结合不同种的细胞,参与免疫应答。
(4)可通过胎盘及粘膜:免疫球蛋白G(IgG)能通过胎盘进入胎儿血流中,使胎儿形成自然被动
免疫。免疫球蛋白A(IgA)可通过及粘膜,是粘膜局部抗感染免疫的主要因素。
(5)具有抗原性:抗体分子是一种蛋白质,也具有刺机体产生免疫应答的性能。不同的免疫球蛋白分子,各具有不同的抗原性。
(6)抗体对理化因子的与一般球蛋白相同:不耐热,60~70℃即被破坏。各种酶及能使蛋白质凝固变性的物质,均能破坏抗体的作用。抗体可被中性盐类沉淀。在生产上常可用硫酸铵或硫酸钠从免疫血清中沉淀出含有抗体的球蛋白,再经透析法将其纯化。
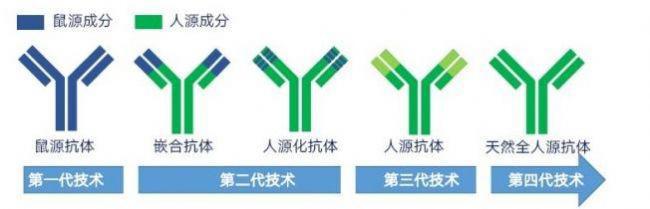
抗体的多样性:
抗体的异质性。抗体的组成极为复杂,是由成千上万、多种多样的免疫球蛋白(Ig)分子所组成。这些Ig分子在形状、大小、结构以及氨基酸的组成和排列上,既相似,又有差别。由于有差别,它们的电泳活性就有很大的变化。
因为抗体具有与抗原决定簇相对应的结合部位(抗原结合簇),所以抗体与抗原的结合具有特异性。另一方面,抗体本身是一种蛋白质,具有本身的氨基酸组成、排列和立体结构,对异种动物来说,它又是抗原。各类Ig都具有可用血清学方法检出的抗原特异性,它们表现出不同的血清学类型。
ARMCX2 ARM重复X连锁蛋白ARMCX2抗体
Apg10 自噬相关蛋白10抗体
ACBD6 酰基辅酶A结合结构域蛋白6抗体
Aminoacylase 1 氨基酰化酶1抗体
Adipose Triglyceride Lipase 脂肪甘油三酯脂酶抗体
phospho-AMPK alpha-1 (Thr172) 磷酸化腺苷单磷酸活化蛋白激酶α1抗体
Amphiregulin 双调蛋白(结肠直肠细胞源性生长因子)抗体
phospho-AQP2(Ser264) 磷酸化水通道蛋白2抗体
AXL 粘附相关激酶抗体
phospho-AXL (Tyr698+Tyr702+Tyr703) 磷酸化粘附相关激酶抗体
phospho-AKT1 + AKT2 + AKT3 (Thr308+Thr309+Thr305) 磷酸化蛋白激酶B抗体
phospho-AQP2(Ser269) 磷酸化水通道蛋白2抗体
phospho-Amyloid Precursor Protein (Thr743) 磷酸化APP淀粉样肽前体蛋白抗体
phospho-AKT1(Ser129) 磷酸化蛋白激酶B抗体
phospho-Amyloid Precursor Protein (Ser198) 磷酸化APP淀粉样肽前体蛋白抗体
phospho-ATF4 (Ser245) 磷酸化活化转录因子4抗体
phospho-Androgen Receptor (Ser213) 磷酸化雄激素受体抗体
phospho-Androgen Receptor (Ser650) 磷酸化雄激素受体抗体
phospho-AKT1+2+3 (Tyr315+316+312) 磷酸化蛋白激酶AKT1,2,3抗体
phospho-AKT1(Thr34) 磷酸化蛋白激酶B抗体
phospho-ATF2(Ser472) 磷酸化活化复制因子2抗体
(1)特异性结合抗原:抗体本身不能直接溶解或杀伤带有特异抗原的靶细胞,通常需要补体或吞噬细胞等共同发挥效应以清除病原微生物或导致。然而,抗体可通过与病毒或毒素的特异性结合,直接发挥中和病毒的作用。
(2)活补体:IgM、IgG1、IgG2和IgG3可通过经典途径激活补体,凝聚的IgA、IgG4和IgE可通过替代途径激活补体。
(3)结合细胞:不同类别的免疫球蛋白,可结合不同种的细胞,参与免疫应答。
(4)可通过胎盘及粘膜:免疫球蛋白G(IgG)能通过胎盘进入胎儿血流中,使胎儿形成自然被动
免疫。免疫球蛋白A(IgA)可通过及粘膜,是粘膜局部抗感染免疫的主要因素。
(5)具有抗原性:抗体分子是一种蛋白质,也具有刺机体产生免疫应答的性能。不同的免疫球蛋白分子,各具有不同的抗原性。
(6)抗体对理化因子的与一般球蛋白相同:不耐热,60~70℃即被破坏。各种酶及能使蛋白质凝固变性的物质,均能破坏抗体的作用。抗体可被中性盐类沉淀。在生产上常可用硫酸铵或硫酸钠从免疫血清中沉淀出含有抗体的球蛋白,再经透析法将其纯化。
抗体的多样性:
抗体的异质性。抗体的组成极为复杂,是由成千上万、多种多样的免疫球蛋白(Ig)分子所组成。这些Ig分子在形状、大小、结构以及氨基酸的组成和排列上,既相似,又有差别。由于有差别,它们的电泳活性就有很大的变化。
因为抗体具有与抗原决定簇相对应的结合部位(抗原结合簇),所以抗体与抗原的结合具有特异性。另一方面,抗体本身是一种蛋白质,具有本身的氨基酸组成、排列和立体结构,对异种动物来说,它又是抗原。各类Ig都具有可用血清学方法检出的抗原特异性,它们表现出不同的血清学类型。
ARMCX2 ARM重复X连锁蛋白ARMCX2抗体
Apg10 自噬相关蛋白10抗体
ACBD6 酰基辅酶A结合结构域蛋白6抗体
Aminoacylase 1 氨基酰化酶1抗体
Adipose Triglyceride Lipase 脂肪甘油三酯脂酶抗体
phospho-AMPK alpha-1 (Thr172) 磷酸化腺苷单磷酸活化蛋白激酶α1抗体
Amphiregulin 双调蛋白(结肠直肠细胞源性生长因子)抗体
phospho-AQP2(Ser264) 磷酸化水通道蛋白2抗体
AXL 粘附相关激酶抗体
phospho-AXL (Tyr698+Tyr702+Tyr703) 磷酸化粘附相关激酶抗体
phospho-AKT1 + AKT2 + AKT3 (Thr308+Thr309+Thr305) 磷酸化蛋白激酶B抗体
phospho-AQP2(Ser269) 磷酸化水通道蛋白2抗体
phospho-Amyloid Precursor Protein (Thr743) 磷酸化APP淀粉样肽前体蛋白抗体
phospho-AKT1(Ser129) 磷酸化蛋白激酶B抗体
phospho-Amyloid Precursor Protein (Ser198) 磷酸化APP淀粉样肽前体蛋白抗体
phospho-ATF4 (Ser245) 磷酸化活化转录因子4抗体
phospho-Androgen Receptor (Ser213) 磷酸化雄激素受体抗体
phospho-Androgen Receptor (Ser650) 磷酸化雄激素受体抗体
phospho-AKT1+2+3 (Tyr315+316+312) 磷酸化蛋白激酶AKT1,2,3抗体
phospho-AKT1(Thr34) 磷酸化蛋白激酶B抗体
phospho-ATF2(Ser472) 磷酸化活化复制因子2抗体